Летальные гены примеры. Рецеccивные, доминантные, летальные гены, мутации

 Live Journal
Live Journal Facebook
Facebook Twitter
TwitterДо сих пор мы рассматривали генетику человека по типам наследования, теперь мы обратимся к одной очень важной категории генов, а именно к летальным и сублетальным генам. Летальные и сублетальные гены представляют собой класс мутаций с заметно выраженным отрицательным влиянием на развитие. Как мы увидим дальше, мутации в очень многих случаях отягощают и разрушают наследственность человека. Это

отягощение мутациями особенно ярко проявляется под действием летальных генов.
Летальные и сублетальные аллели могут быть доминантными и рецессивными. В случае доминантного леталя ребенок погибает во внутриутробном периоде или в раннем младенческом возрасте. Вполне понятно, что такие гены не могут передаваться по наследству. Их наличие говорит о появлении мутаций. Суммарная частота таких доминантных леталей не так мала. Особенно их количество велико при действии радиации. В генетике мышей и дрозофилы при анализе зависимости частоты мутаций от дозы и при анализе других особенностей мутагенных воздействий широко используются доминантные летали. При наличии сублеталей, иначе называемых полулетальными генами, ребенок умирает в раннем детстве или несколько позднее, но, как правило, до достижения половой зрелости. Примером доминантной сублетальной мутации может служить эпилойя - заболевание, характеризующееся действием целого синдрома отрицательных признаков. Дети, больные эпилойей, имеют патологические разрастания кожи, им свойственна умственная отсталость, эпилепсия, опухоли сердца, почек и других органов. В редких случаях больные эпилойей выживают и вступают в брак со здоровым супругом. Половина детей от такого брака оказывается больными врожденной эпилойей (рис. 131).
Другим ярким примером сублетальной доминантной мутации может послужить заболевание ретинобластома. Этот


доминантный ген вызывает раковую опухоль в глазу, что почти всегда ведет к смерти ребенка в раннем возрасте.
Однако основная масса врожденных уродов падает на рецессивные летали и сублетали. В этом случае рецессивные вредные гены скрываются в генотипе внешне здоровых родителей, однако, когда две таких гетерозиготы вступают в брак, среди их детей около 25% оказываются больными.
На рисунке 132 представлены основные типы скрещиваний в популяциях людей, которые осуществляются при наследовании рецессивных летальных и сублетальных генов. В квадрате 1 представлен тип браков между генетически нормальными людьми. У обоих родителей и у всех их потомков все аллели представлены нормальными генами (аллель +). В квадрате 2 изображен брак между генетически здоровым человеком и гетерозиготой (символ а). В этом случае отрицательный аллель передается половине потомства, оставаясь, однако, при этом в скрытом виде. В квадрате 3 генетически здоровый человек вступил в брак с больным (символ аа), все дети внешне здоровы, но в гетерозиготном состоянии несут сублетальную мутацию а. В квадрате 4 гетерозигота вступает в брак с больным; половина их детей внешне здорова, но несет сублетальную мутацию, другая половина больные. Наконец, квадрат 5 при браке двух больных (символы аа) все потомство будет больным. В качестве врожденных болезней, вызываемых действием рецессивных летальных и сублетальных генов, можно назвать детскую амавротическую идиотию, при которой мозговая ткань дегенерирует, наступает слепота и ребенок гибнет; врожденную патологию кожи с наличием
глубоких кровоточащих трещин - ихтиоз (рис. 133); врожденный детский паралич и целый ряд других врожденных заболеваний.
Многие ранние самопроизвольные выкидыши вызываются выщепляющимися летальными генотипами.
Известны доминантные мутации, в гетерозиготах определяющие какой-либо более или менее нейтральный признак, которые, однако, в гомозиготном состоянии оказывают летальное действие. Так, например, аллель пигментации ксеродермы в гетерозиготном состоянии вызывает сильную веснушчатость. Однако у гомозигот развивается пигментная ксеродерма. Это сублетальное заболевание (рис. 134), при котором действие света вызывает поражение кожи, приводящее затем к злокачественному росту.
Один из аллелей у человека в гетерозиготном состоянии вызывает избыточное содержание холестерина в крови. Однако дети, гомозиготные по этому аллелю, заболевают тяжелой сублетальной аномалией.
Во всех этих случаях мы имеем классическую картину наследования доминантов с рецессивным летальным эффектом, ранее установленную в опытах с дрозофилой, мышами и другими организмами. Например, у мыши хорошо известен аллель A Y , который у гетерозигот вызывает желтую окраску шкурки. Однако все попытки получить чистую расу желтых мышей остались безуспешными. При скрещивании двух желтых мышей в среднем всегда на две желтые мыши рождается одна другой окраски, например черная. Природа этого явления нашла свое объяснение в том, что ген доминантной желтой окраски оказался
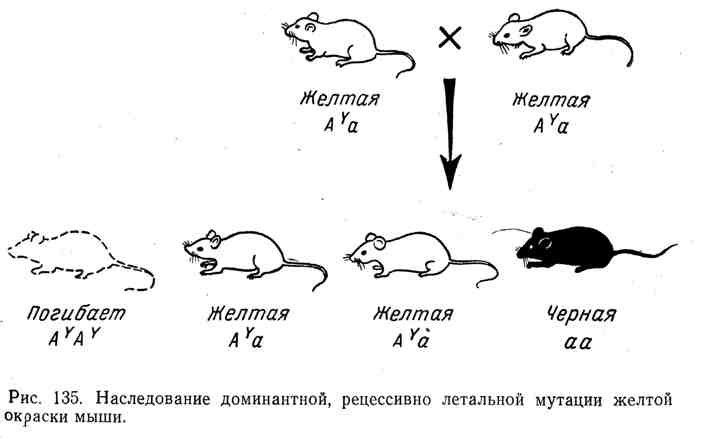
рецессивным леталем. Все гомозиготы ( A Y A Y ) погибали. Желтая мышь всегда гетерозиготна, наряду с аллелем A Y она содержит другой аллель, например а (черной окраски). Ход наследования при скрещивании гетерозигот A Y a и расщепления по формуле 2: 1 изображены на рисунке 135.
— Источник—
Дубинин, Н.П. Горизонты генетики/ Н.П. Дубинин. – М.: Просвещение, 1970.- 560 с.
Post Views: 121
Почему один ген является доминантным, а другой рецессивным? Мутантная ДНК, не кодирующая никакой белок или кодирующая белок, лишенный активности, не будет эксп-рессироваться, т.е. представляет собой рецессивный ген. У гетерозиготных индивидуумов нормальный ген направляет синтез нормального белка, а рецессивный ген никакого вклада в синтез функционального белка не вносит. Такой индивидуум имеет нормальный фенотип; нормальный ген в этом случае доминирует. У индивидуума, гомозиготного по данному рецессивному гену, соответствующий белок не образуется, а поэтому нормальная экспрессия данного признака невозможна. Рецессивный фенотип - это отсутствие нормального признака. Так, карликовость - результат «отсутствия ростовых веществ».
Если белок, о котором идет речь, необходим для жизни данного организма, то организм, неспособный образовать активную форму этого белка, обречен на гибель, и дефектный ген называется в этом случае летальным геном. Доминантные летальные гены существуют, но в большинстве случаев они быстро элиминируются, потому что вызывают гибель несущего их организма. Рецессивные же летальные гены могут и не причинять вреда гетерозиготному индивидууму, поэтому они могут передаваться следующим поколениям и распространяться в популяции весьма широко. Подсчитано, что в среднем человек гетерозиготен приблизительно по 30 летальным рецессивным генам. Это больше, чем у многих других организмов, и отчасти именно этим объясняется тот факт, что у людей среди потомков от браков между близкими родственниками процент летальных наследственных признаков выше, нежели при близкородственных скрещиваниях у большинства других видов.
Если одна копия «нормального» гена не обеспечивает образования его белкового продукта в количестве, достаточном для нормального функционирования организма, то по данному гену наблюдается неполное доминирование. В таких случаях гетерозигота отличается по фенотипу от обеих гомозигот. У человека, например, имеется летальный ген, обусловливающий в гетерозиготном состоянии брахидактилию, т.е. укорочение средней фаланги пальцев; палец при этом выглядит так, как будто в нем не три, а две фаланги. У гомозигот этот ген является причиной аномального развития скелета. Дети, гомозиготные по этому гену, рождаются без пальцев и с другими нарушениями в развитии скелета, приводящими к смерти в раннем возрасте.
В тех случаях, когда брахидактилией страдают оба родителя, можно ожидать, что из каждых четырех детей один будет гомозиготен по этому летальному гену и умрет в раннем детстве, два будут гетерозиготны, т.е. будут страдать брахи-дактилией, и один будет нормальным.
Широко известный ген человека, который в гомозиготном состоянии часто оказывается летальным, ответствен за серповидноклеточную анемию. Соответствующий нормальный ген кодирует одну из полипептидных цепей (бета-цепь) гемоглобина-белка, функция которого заключается в переносе кислорода; этот.белок находится в эритроцитах и обусловливает их красный цвет. У мутантного гена один из нуклеотидов, присутствующих в нормальной ДНК, заменен другим, в результате чего и в бета-цепи гемоглобина одна из аминокислот заменена другой.
Это изменение, казалось бы столь ничтожное, влечет за собой роковые последствия. При аномально низком содержании кислорода молекулы гемоглобина в эритроцитах, содержащих гемоглобин S, агрегируют, образуя волокна. Вследствие этого эритроциты деформируются, приобретая серповидную форму. Такие эритроциты застревают в мелких кровеносных сосудах, и кровоснабжение участков, обслуживаемых этими сосудами, нарушается. Нарушение кровоснабжения вызывает разнообразные симптомы: чувство усталости, судороги, раздражительность.
Ген серповидноклеточности и соответствующий нормальный ген кодоминантны, так что у гетерозиготных индивидуумов синтезируются и нормальные и мутантные бета-цепи. Эритроциты таких людей приобретают серповидную форму лишь при крайне низком содержании кислорода, например, на больших высотах. Без специальных тестов такие гетерозиготные индивидуумы могут и не знать о том, что они являются носителями гена серповидноклеточности. Гомозиготные индивидуумы страдают гораздо сильнее, потому что у них дефектны все бета-цепи.
Индивидуумов, гетерозиготных по тому или иному генетическому нарушению, называют носителями, а гомозиготных- больными. Про людей, гетерозиготных по гену серповидноклеточности, иногда говорят, что они больны серповидноклеточной анемией. Это неверно, ибо это предполагает, что носитель данного гена менее жизнеспособен, нежели нормальный гомозиготный индивидуум. Между тем в действительности это обычно не так.
Около 90% индивидуумов, гомозиготных по гену серповиднооклеточности, умирает в раннем возрасте. Можно было бы ожидать, что такой летальный ген будет быстро элиминироваться естественным отбором, поскольку гомозиготы по этому гену умирают слишком рано и не могут передать его потомству. Однако в тропической Африке на весьма обширных пространствах от 20 до 40% людей гетерозиготны по этому гену. В пятидесятых годах исследователи обратили внимание на один примечательный факт: оказалось, что эти люди живут в местах где распространена особо вирулентная форма малярии.
Болезнь Тея-Сакса, проявляющаяся в нарушении развития головного мозга и приводящая к смерти в возрасте около четырех лет, обусловлена наличием рецессивного летального гена У индивидуумов, гомозиготных по этому гену, отсутствует фермент; ответственный за метаболические превращения определенного липида в клетках мозга. Без этого фермента липид накапливается, и клетки утрачивают способность нормально функционировать. Лечить эту болезнь пока не умеют. Известно, что среди еврейского населения Восточной Европы из каждых 30 человек один гетерозиготен по данному гену. Однако в США приблизительно одна треть всех случаев болезни Тея-Сакса приходится на долю детей из нееврейских семей.
Среди белого населения США чаще всего встречается летальный ген, обусловливающий кистозный фиброз - болезнь, при которой бронхиолы, атакже протоки поджелудочной железы и печени заполняются густой слизью. Жертвы кистозного фиброза умирают обычно от инфекционных респираторных заболеваний в детском и юношеском возрасте, не доживая до 20 лет. Предполагают, что ген, ответственный за кистозный фиброз, кодирует один из ферментов цепи переноса электронов.
Нарушения расщепления в соотношении 3:1 может быть обусловлено с наличием летальных генов. Гены, вызывающие нарушения в развитии организма называются летальными генами . Одни летальные гены обуславливают крупные аномалии (аборты у животных, рождение мертвого потомства, рождение уродов), другие вызывают нарушение физиологических процессов, третьи – гибель организма на разных стадиях онтогенеза. Сейчас у крупного рогатого скота обнаружено 46 летальных признаков, у лошадей – 10, свиней – 18, овец – 15, кур – 45, индеек – 6, уток – 3, наследование которых хорошо изучено. Всего известно 143 летальных признака. К летальным признакам у крупного рогатого скота относится карликовость, укорочение нижних конечностей, бесшерстность, бульдогообразность, врожденная водянка головного мозга, паралич тазовых конечностей и другие. У свиней – отсутствие конечностей, волчья пасть, отсутствие участков кожи и другие аномалии.
В большинстве случаев летальные гены рецессивны, поэтому долгое время они могут находиться в скрытом состоянии. И гетерозиготные их носители совершенно здоровы и не отличаются от нормальных организмов. Необходимо отметить, что встречаются и доминантные летальные гены, однако они вызывают гибель организма на ранних стадиях онтогенеза. Примером этому служит серебристо-серая окраска смушка у каракульской породы овец. Серебристо-серая окраска смушка на мировом рынке дороже, чем черного каракуля. Серая окраска смушка является доминантным признаком, черная – рецессивным. При скрещивании серых овец с серыми баранами в потомстве появилось 25% черных ягнят, потому что родители были гетерозиготны поэтому признаку. Было установлено, что серых гомозиготных овец нет. Схема наследования серой масти у овец следующая:
Р♀ Сс х ♂ Сс серые
гаметы С с С с
F 1 СС Сс Сс сс
25% 50% серых 25% черных
Однако в дальнейшем было установлено, что 25% ягнят с переходом на пастбище погибла от хронического тимпанита. Причина гибели – нарушения в деятельности парасимпатической нервной системы. Оказалось, что ген серой окраски в гомозиготном состоянии обладал летальным действием. Для прекращения гибели ягнят генетики Васин, Глембоцкий, Михловский предложили спаривать серых овец с черными баранами, где получали 50% серых и 50% черных ягнят и среди серых ягнят уже не наблюдалась гибель.
серые черные
♀ Сс х ♂сс
Сс Сс сс сс
50% серых 50% черных
Аналогичные примеры можно привести с зеркальным карпом.
♀ Аа х ♂Аа - линейный
F 1 АА Аа Аа аа
голый линейный чешуйчатый
Таким образом, летальные гены проявляют свое действие только в гомозиготном состоянии. А в гомозиготное состояние летальные гены чаще всего переходят при родственном спаривании. Если установлено, что родители являются носителями летальных генов в гетерозиготном состоянии, то матку необходимо немедленно выбраковать, а производителя использовать на не родственных ему животных. Летальные гены могут мигрировать из стада в стадо, из одного региона в другой, из одного государства в другое, так как при искусственном осеменении крупного рогатого скота сперму можно транспортировать в любой регион земного шара, и она может храниться в течение 70 лет при t= -196 º С.
Примером действия летального гена может служить бык-производитель Принц-Адольф голштинской породы, который был завезен из Голландии в Швецию, как выдающийся производитель, широкого использовался во многих стадах Швеции. Первое поколение, полученное от этого быка-производителя, было нормальным, а при дальнейшем спаривании начали рождаться бесшерстные телята. Значит, бык-производитель Принц-Адольф был гетерозиготен по рецессивному летальному гену. И видимо половина его дочерей, внучек, правнучек унаследовали этот летальный ген бесшерстности. При родственном спаривании этого быка-производителя с дочерями, внучками и правнучками летальный ген переходил в гомозиготное состояние и вызвал появление бесшерстных телят.
Таким образом, зная закономерности наследования летальных генов, можно успешно вести борьбу с вредными их последствиями.
В селекции растений и животных, а также в научных исследованиях часто применяют возвратное и анализирующее скрещивание. Сущность возвратного скрещивания состоит в том, что гибриды первого поколения скрещиваются с родительскими формами.
Например, схема:
I ♀ КК х ♂ кк II ♀ Кк х ♂ КК III ♀ Кк х ♂ кк
F 1 Кк Кк Кк Кк F 2 КК Кк КК Кк F Кк Кк кк кк
комолые комолые комолые рогатые
Ф 1: 0 Ф 1: 0 КК - 2 Ф 1: 1
Г 1: 0 Г 1: 1 Кк - 2 Г 1: 1
Для определения состава гамет, данная особь является гомозиготной или гетерозиготной по некоторым генам, для этого применяют анализирующее скрещивание. При этом испытываемая особь, имеющая доминантный признак, скрещивается с гомозиготной особью, имеющий рецессивный признак.
Скрещивание особей с рецессивной родительской формой получило название анализирующего. Анализирующее скрещивание широко применяется при гибридологическом анализе, когда нужно установить генотип интересующей нас особи.
Схема скрещивания:
АА Р. ♀ АА х ♂аа ♀ Аа х ♂ аа
гаметы А А а а А а
Аа черная масть F 1 Аа Аа Аа Аа F Аа Аа аа аа
черные черная красная
Если все особи будут иметь доминантный признак, исследуемая особь будет гомозиготна, а если появится рецессивный признак, она гетерозиготна.
Парные контрастные признаки, дающие при скрещивании расщепления 3:1 называются аллеломорфными признаками, а гены, обуславливающие их развитие аллелями.
Аллели – форма состояния гена. И ген находится в двух аллельных состояниях – доминантные и рецессивные аллели. Аллели – это парные гены, которые локализованы в одинаковых локусах парных гомологичных хромосом и определяют направления развития признака.
Множественным аллелизмом называют различное состояние одного и того же гена обусловленного мутациями. Кролики – окраска черная СС, шиншилла, гималайский альбинос. Муха дрозофила, ген уайт, окраска глаз красная 20 аллелей. Крупный рогатый скот в группе «В» 300 аллелей.
Управление доминированием
После открытия законов или правил доминирования долгое время считали, что доминирование и рецессивность определяются наследственными факторами (генами) и не зависят от условий внешней среды. Однако, работами И.В. Мичурина было доказано, что доминирование зависит не только от наследственных факторов, но и от условий внешней среды, в которых развивается организм. В своих опытах он показал, что при скрещивании местных сортов растений с южными сортами, т.е. завезенных с других зон, регионов, доминируют признаки местного сорта.
Большая заслуга И.В. Мичурина состоит в том, что он разработал учение об управлении доминированием. Он показал пути, какими можно изменить доминирование в нужном направлении. Если гибридам создать условия близкие к одному из родителей, то у гибридов будут развиваться признаки этого родителя. Пример из животноводства, местные киргизские лошади скрещивались с чистокровной верховой. В зависимости как будет выращиваться потомство, такие будут получены результаты. Учение об управлении доминированием имеет большое значение в практике животноводства. Создавая животным, различные условия внешней среды, тем самым регулируем развитие желательных признаков. Молочный тип – кормление грубыми и сочными кормами, моцион, подготовка к отелу, массаж вымени, раздой первотелок и другие мероприятия. Мясной тип – кормление концентрированными кормами, уменьшение количества прогулок, привязное содержание и другие мероприятия. Таким образом, управление доминированием имеет большое значение в практике животноводства.
Стадиях развития, но существуют летали, вызывающие гибель например, при окукливании личинки дрозофилы). Летальные аллели возникают в результате т. н. летальных мутаций - летальность таких мутаций говорит о том, что данный ген ответственен за какую-либо жизненно необходимую функцию.
Летальными называются аллели, носители которых погибают из-за нарушений развития или заболеваний, связанных с работой данного гена. Между летальными аллелями и аллелями, вызывающими наследственные болезни, есть все переходы. Например, больные хореей Хантингтона (аутосомно-доминантный признак) обычно умирают в течение 15-20 лет после начала заболевания от осложнений, и в некоторых источниках предлагается считать этот ген летальным.
Сублетальными, или полулетальными называются аллели, эффект гибельности которых част, но не обязателен (то есть переходные между летальными аллелями и аллелями, вызывающими наследственные болезни), условно летальными называют мутации, при которых организм несущий такие мутации может жить в предельно узком диапазоне условий, например мутации ауксотрофности у микроорганизмов (не способность расти на питательных средах без определённых жизненнонеобходимых веществ из-за утраты способности их синтезировать) субстратнозависимые мутации (неспособность использовать некоторые вещества в качестве источника углерода и энергии) и температурнозависимые мутации (способность жить только в узком диапазоне температур - например некоторые мутанты дрозофилы не способны жить при температуре выше 25 о С).
Примечания
Wikimedia Foundation . 2010 .
Смотреть что такое "Летальные гены" в других словарях:
ЛЕТАЛЬНЫЕ ГЕНЫ - ЛЕТАЛЬНЫЕ ГЕНЫ, гены (см.), вызывающие смертельный исход в гомозиготном состоянии. Наряду с ними известно большое число полулетальных факторов, приводящих очень часто к рождению различного рода нежизнеспособных уродов или просто тем или иным… …
Летальные гены - * лятальныя гены * lethal genes …
гены синтезирующая машина - * машына, якая сінтэзуе гены * gene machine автоматический ДНК синтезатор для получения коротких (как правило, длиной в 15 30 п. о.) нитей ДНК для использования их в полимеразной цепной реакции. Гены сложные * гены складаныя * compound genes гены … Генетика. Энциклопедический словарь
Летальные факторы - * лятальныя фактары * lethal factors менделирующие () единицы (гены и хромосомные аберрации), обусловливающие гибель (летальный исход) организма до достижения им половой зрелости. Л. ф. классифицируют: а) по степени пенетрантности (см.); б) по… … Генетика. Энциклопедический словарь
ЛЕТАЛЬНЫЕ ФАКТОРЫ - Менделирующие единицы (гены, хромосомы), обусловливающие гибель организма ранее достижения им половой зрелости. Летальные факторы классифицируются: 1. По степени гибели организмов: абсолютные, приводящие к гибели 100% особей; сублетальные более… … Термины и определения, используемые в селекции, генетике и воспроизводстве сельскохозяйственных животных
ЧЕЛОВЕК - ЧЕЛОВЕК. Содержание: Происхождение человека............ 5 24 Наследственность человека........... 530 Человек по систематическому положению в органическом мире относится к царству животных (Animalia), к полцарству многоклеточных (Metazoa), к… … Большая медицинская энциклопедия
НАСЛЕДСТВЕННОСТЬ - НАСЛЕДСТВЕННОСТЬ, явление передачи потомству материальных факторов, определяющих развитие признаков организма в конкретных условиях среды. Задачей изучения Н. является установление закономерностей в возникновении, свойствах, передаче и… … Большая медицинская энциклопедия
ГЕНЕТИКА - (от греч. genesis происхождение), обычно определяется как физиология изменчивости и наследственности. Именно так определил содержание генетики Бетсон (Bateson), предложивший в 1906 г. этот термин, желая подчеркнуть, что из трех основных элементов … Большая медицинская энциклопедия
ГЕН - (от греч. gignomai становлюсь), наследственный задаток; термин введен Иогансеном (Johannsen) с расчетом, чтобы он не содержал в себе «никакой гипотезы», и противопоставлен им понятию «наследственный признак». Находясь в… … Большая медицинская энциклопедия
Эта статья о биологическом таксоне. Об обиходном понятии см. Гриб. Грибы … Википедия
Изменение расщепления по фенотипу в отношении 3:1 во втором поколении моногибридного скрещивания бывает связано с разной жизнеспособностью зигот Ф2. Разная жизнеспособность зигот мэжет быть обусловлена наличием летальных генов. Летальным называется ген, вызывающий нарушения в развитии организма, что приводит его к гибели или уродству.
Изучение врожденных аномалий показало, что при разных летальных генах гибель особей бывает различной и может происходить на разных стадиях развития.
По классификации, предложенной Розенбауэром (1969), гены, вызывающие гибель 100 % особей до достижения ими пЪловой зрелости, называются летальными, более 50 % -сублетальными (полулетальными) и менее 50 % - субвитальными. Однако следует отметить, что разделение это до некоторой степени условно и иногда не имеет четких границ. Примером может служить сцепленная с полом голость у кур. Почти половина голых цыплят гибнет в последние 2-3 дня инкубации. Из числа вылупившихся около половины цыплят гибнет до 6-недельного возраста, если их выращивают при температуре 32-35 "С. Но если температура в брудерах будет повышена на 5,5 °С, то погибнет значительно меньше голых цыплят. В 4-5 мес у голых цыплят вырастает редкое оперение и они уже в состоянии переносить довольно низкие температуры. В естественных условиях эта мутация, по-видимому, будет летальной и приведет к 100%-ной гибели птиц. Приведенный пример показывает, что характер проявления полулетального гена в значительной мере может зависеть от условий среды.
Летальные гены могут быть доминантными и рецессивными. В числе первых летальных факторов был открыт аллель, обусловливающий желтую окраску мышей. Ген желтой окраски является доминантным (Y). Его рецессивный аллель (у) в гомозиготном состоянии обусловливает появление черной окраски. Скрещивание желтых мышей между собой давало в потомстве две части особей желтых и одну часть черных, т. е. получалось расщепление 2:1, а не 3:1, как следовало из правила Менделя. Оказалось, что все взрослые мыши гетерозиготны (Yy). При скрещивании между собой они должны были дать одну часть гомозиготного потомства по желтой окраске (IT), но оно погибает еще в эмбриональном периоде, две части гетерозигот (Yy) будут желтыми и одна часть гомозигот по рецессивному признаку (уу) будет черной. Схема скрещивания выглядит так:
Таким же образом наследуется серая окраска шерсти у каракульских овец (сокольских, малич и др.), платиновая окраска у лисиц, распределение чешуи у линейных карпов и т. д.
Летальные гены в большинстве случаев рецессивны и поэтому долгое время могут находиться в скрытом состоянии. Совершенно здоровое и нормальное по фенотипу животное может быть носителем летального гена, действие которого обнаруживается только при переходе в гомозиготное состояние. В гомозиготное состояние летальные гены чаще всего переходят при родственном спаривании. В практике животноводства при разведении лошадей был случай гибели 25 жеребят на 2-4-й день после рождения от уродства прямой кишки - отсутствия заднепроходного отверстия (Atresia ani). Оказалось, что все жеребцы и кобылы, от которых рождались такие ненормальные жеребята, происходили от одного жеребца. Он был гетерозиготен по летальному гену (LI). Вначале этот жеребец при скрещивании с нормальными кобылами (LL) дал потомство, по фенотипу нормальное, но по генотипу половина потомков была благополучной (LL), a половина - гетерозиготной (LI), несущей рецессивный задаток (0 летального гена. При родственном спаривании гетерозиготных животных (Ы х Ы) появилась часть жеребят, гомозиготных по летальному гену (II), с уродством прямой кишки. Они все погибли. (Более подробно об аномалиях при летальных генах будет сказано в соответствующей главе.)
Конец работы -
Эта тема принадлежит разделу:
Предмет и методы генетики
Виды наследственности.. клетка как генетическая.. митоз..
Если Вам нужно дополнительный материал на эту тему, или Вы не нашли то, что искали, рекомендуем воспользоваться поиском по нашей базе работ:
Что будем делать с полученным материалом:
Если этот материал оказался полезным ля Вас, Вы можете сохранить его на свою страничку в социальных сетях:
| Твитнуть |
Все темы данного раздела:
Методы генетики
Специфические методы генетики.
1. Гибридологический метод (открытый Менделем). Основные черты метода:
а). Мендель учитывал не весь многообразный комплекс признаков у родите
Предмет генетики
Наследственность - это воспроизведение жизни (Н. П. Дубинин). Изменчивость - это возникновение различии между организмами по ряду признаков и свойств.
Наследственность, изменчивос
Виды изменчивости
Выделяют следующие виды изменчивости: мутационную, ком-бинативную, коррелятивную и модификационную.
Мутационная изменчивость. Мутация - стойкое изменение в структуре ДНК и кариотипе
Роль ядра и других органелл в прередаче, сохранения и реализации наследственной информации
Организм животных состоит из миллиардов клеток, и все они ведут начало от двух половых клеток (гамет) - отцовского спер-мия и материнской яйцеклетки. При слиянии гамет - о
Роль ядра и цитоплазмы в наследственности
Ядро - основной компонент клетки, несущий генетическую информацию. Оно может находиться в двух состояниях: покоя - интерфазы и деления - митоза или мейоза. Интерфазное ядро представляет собой кругл
Морфологическое строение и химический состав хромосом
При микроскопическом анализе хромосом прежде всего видны различия их по форме и величине. Строение каждой хромосомы сугубо индивидуальное. Можно заметить также, что хромосомы обладают общими морфол
Кариотип и его видовые ообенности
Анализ хромосом в клетках животных и растений разных видов позволил выявить ряд общих закономерностей, имеющих важное значение при изучении явлений наследственности и изменчивости. Установлено, чт
Роль генотипа и условий среды в формировании фенотипа
Фенотип каждого организма формируется под влиянием генотипа и условий среды. Генотип определяет норму реакции организма - границы изменчивости выражения признака под влиянием и
Особенности гаметогенеза самцов и самок
Процесс развития половых клеток носит название гаметогене-за. У самцов этот процесс называется спермиогенезом, а у самок - овогенезом (рис. б). Половые клетки в развитии после
Полиплоидия и ее значения
Полиплоидия (тут- «полп» -^* много, «плоид» -*- складывать) увеличение числа хромосом, кратное (эуплоидия) и некратное
(анэуплоидия) гаплойдному набору: 3н - триплоид 4н тетраплоид.
Паталогии мейоза и митоза и ее значения
Патология мейоза.Основная патология мейоза - нерасхождение хромосом. Оно может быть первичным, вторичным и третичным. Первичное нерасхождение возникает у особей с нормальным кар
Оплодотворение
Оплодотворение- процесс слияния яйцеклетки со сперматозоидом. Яйцеклетка^- женская гамета (половая клетка) -*- ужи-вотных образуется в ядониках. Она формируется в результате оогенеза и соде
Особенности гибридологического метода менделя
Сущность гибридологического метода заключается в следующем:
1) для скрещивания выбирают родительские формы, четко
различающиеся по одной, двум или трем парам контрастных,
Закон единообразия гибридов первого поколения
Мендель избрал для анализа семь пар четко различающихся признаков: форма семян - круглая или морщинистая, окраска семядолей - желтая или зеленая, окраска цветков - пурпурная или белая, рост растени
Закон расщепления
В следующем опыте путем скрещивания между собой гибридов первого поколения Мендель получил второе поколение, в котором наряду с доминирующими признаками проявились и рецессивные. Появление во втор
Аллели. Множественный аллелизм
Аллельными генами (аллелями) называют гены, расположенные в одинаковых точках (локусах) парных гомологичных хромосом. Аллели оказывают влияние на развитие одного и того же признака организм
Анализирующее скрещивание. Правило чистоты гамет
Мендель провел опыт по скрещиванию гибридов первого поколения с растениями гороха исходных родительских сортов. Скрещивание гибридов первого поколения (Аа) с особями, сходными по генотипу
Полигибридное скрещивание
Положение о независимом наследовании разных пар аллелей и признаков было подтверждено Менделем при изучении наследования трёх пар признаков у гороха. Он скрещивал сорт растения с круглыми семенам
Плейотропное действие генов
Плейотропия.Это влияние одного гена на развитие двух и более признаков (множественное действие гена). Так, Д. К. Беляев и А. И. Железнова установили, что у норок большинство мутац
Полимирия
Иногда на формирование признака влияют две или несколько пар неаллельных генов. Проявление признака в этом случае зависит от характера их взаимодействия в процессе развития организма. В первом по
Сцепленное наследование признаков
Мы познакомились с дигибридным скрещиванием и уяснили, что независимое комбинирование признаков объясняется тем, что расщепление одной пары аллельных генов, определяющих соответствующие признаки, п
Полное сцепление
Т. Морган скрещивал черных длиннокрылых самок с серыми с зачаточными крыльями самцами. У дрозофилы серая окраска тела доминирует над черной, длиннокрылость - над зачаточными крыльями. Обозначим ге
Неполное сцепление
В следующем опыте, так же как и в предыдущем, Морган скрещивал черных длиннокрылых самок с серыми зачаточно-крылыми самцами. В первом поколении получил все потомство серое длиннокрылое. Затем снова
Картирование хромосом
Сущность соматического кроссинговера заключается в том, что он осуществляется при митотическом делении соматических клеток главным образом эмбриональных тканей. Кроссинговер происходит меж
Карты хромосом
После того как была установлена связь генов с хромосомами и обнаружено, что частота кроссинговера всегда вполне определенная для каждой пары генов, расположенных в одной группе сцепления, встал во
Гомо и гетерогаметный пол
В ходе эволюции у большинства раздельнополых организмов сформировался механизм детерминации пола, обеспечивающий образование равного количества самцов и самок, что необходимо для
Наследование признаков, ограниченных полом
Наследование признаков, контролируемых генами, локализованными в аутосомах, но фенотипически проявляющихся исключительно или преимущественно у одного пола, - есть наследование, ограниченное п
Проблема регуляции пола
Проблема регуляции пола вытекает из необходимости увеличения продукции животноводства за счет преимущественного получения особей одного вида, дающих более высокий выход молока, мяса, шерсти, яиц и
Наследование признаков, сцепленных с полом
Причины более высокой смертности среди самцов млекопитающих можно объяснить исходя из особенностей наследования признаков, сцепленных с полом. Явление сцепленного с полом наследования впервые откр
Нуклеиновые кислоты, доказательства их роли в наслндственности
Генетическая информация реализуется в процессе биосинтеза белков. Все основные свойства живых существ определяются структурой и функцией белковых молекул. В последние 40 лет в ряде лаборатор
Химический состав и структура нуклеиновых кислот
Нуклеиновые кислоты впервые открыл И. Ф. Мишер в 1868 г. Он выделил из ядер клеток особое вещество кислотной природы и назвал его нуклеином. Впоследствии ему дали название «нуклеиновая кислота». Б
Днк рнк
Пуриновые основания Аденин, гуанин Аденин, гуанин
Пиримидиновые основания Цитозин, тимин Цитозин, урацил
Углеводный компонент Дезоксирибоза Рибоза
Об
Строение и типы рнк
Многочисленными исследованиями было установлено, что синтез белка в клетке происходит не в ядре, где находится ДНК, а в цитоплазме. Следовательно, сама ДНК не может служить матрицей для синтеза бе
Репликация ДНК
Репликация (удвоение) ДНК.ДНК находится в хромосомах, и репликация ее происходит перед каждым удвоением хромосом и делением клетки. Дж. Уотсон и Ф. Крик предложили схему уд
Генетический код и его свойства
Представление о том, что генетическая информация о структуре белковых молекул зашифрована в ДНК путем определенного расположения нуклеотидов, конкретизировал Ф. Крик в гипотезе последовательност
Синтез белка в клетке
В настоящее время можно считать установленным, что наследственность реализуется в процессе биосинтеза белка. Синтез ферментов и других белков, необходимых для жизнедеятельности и развития организм
Конъюгация
Конъюгация - перенос генетического материала от одной бактериальной клетки (донора) к другой (реципиенту) при их непосредственном контакте. Процесс конъюгации у бактерий обнаружили Дж. Ле
Трансдукция
Трансдукция - перенос генов из одной бактериальной клетки в другую при помощи бактериофага. Впервые это явление установили в 1952 г. Н. Зиндер и Дж. Ледерберг. Они проводили исследования на
Трансформация
Трансформация - поглощение изолированной ДНК бактерии донора клетками бактерии реципиента. Явление трансформации кратко освещено при изложении доказательств роли ДНК в наследственно
Мутационная изменчивость
В предыдущих главах отмечено одно из основных свойств кариотипа, ДНК и ее участков (генов) - сохранять постоянство внешнего и внутреннего строения. Морфофункциональная устойчивость генетического м
Хромосомные мутации
Изменения кариотипа могут быть количественными, структурными и одновременно теми и другими. Рассмотрим отдельные формы изменения хромосом (см. схему).
Числовые мутации кариотипа.
Генные мутации
По характеру действия генные мутации могут быть доминантными или рецессивными. Чаще мутантный ген обладает рецессивным эффектом. Нормальный аллель подавляет при этом действие измен
Регуляция синтеза ирнк и белка
Все клетки организма, как бы они ни были дифференцированы, как правило, тождественны по генотипу. В каждой клетке "имеются все хромосомы и весь набор свойственных данному организму генов. Однако к
Строение и размножение бактерий
Химический состав клеток бактерий в основном такой же, как и клеток высокоорганизованных организмов. Клетки бактерий окружены оболочкой, внутри которой находятся цитоплазма, ядерный аппарат, рибосо
Строение и размножение вирусов
Вирусы относятся к микроорганизмам, хотя резко отличаются от всех известных клеточных форм жизни. Частицы вирусов очень малы (от 20 до 450 нм). С помощью электронного микроскопа обнаружено, что он
Взаимодействие фага с бактериальной клеткой
К клеточной стенке бактерий фаги прикрепляются концевыми нитями отростков. Затем оболочка бактерии растворяется с помощью фермента лизоцима, белковый чехол хвостового отростка сокращается и через
Анеуплоидия
Числовые аномалии кариотипа (анеуплоидия). Числовые аномалии хромосом относят к вновь возникающим мутациям. Однако имеются исследования, которые показывают, что может быть семейная предрасположенно
Транслокации
Структурные мутации хромосом. Транслокации. Наибольшее количество исследований у крупного рогатого скота проведено по изучению частоты и влияния на плодовитость центрического слияния - тран
Генетические анамалии у с.х. животных
У сельскохозяйственных животных известны десятки аномалий, возникновение которых связано с рецессивными или доминантными мутациями генов. Эти аномалии встречаются в отдельных популяциях с разной
Простой аутосомный рецессивный тип наследования
Этим термином пользуются, когда аномалия обусловлена одним рецессивным геном, локализованным в аутосоме. Иногда употребляют термин моногенно-аутосомный тип наследования, что является синонимом. Ано
Аутосомный доминантный тип наследования
Признаки, обусловленные доминантными генами, как правило, проявляются в гетерозиготном состоянии. Возможный вариант скрещивания и характер расщепления следующие (табл. 40). При таком доминантном
Сцепленный с х-хромосомой тип наследования
В дополнение к главе «Генетика пола» отметим следующее. Гены, локализованные в Х-хромосоме, могут проявлять доминантный или рецессивный эффект. Возможные варианты скрещиваний и расщеплений при сц
Мультифакториальное наследование
Развитие исследований в генетике, в том числе и ветеринарной, привело к выводам о том, что далеко не при всех формах или даже случаях патология - простой менделирующий признак. Часто она обусловле
Практическое значение закона
ПОПУЛЯЦИЯ И «ЧИСТАЯ ЛИНИЯ»
По Н. В. Тимофееву-Ресовскому, популяция - это совокупность особей данного вида, в течение длительного времени (большого числа поколений
Структура свободно размножающейся популяции. Закон харди - вайнберга
Харда" и Вайнберг провели математический анализ распределения генов в больших популяциях, где нет отбора, мутаций и смешивания популяций. Они установили, что такая популяция находится в состоянии
Генная инженерия
Генная инженерия - раздел биотехнологии, связанный с целенаправленным конструированием in vitro новых комбинаций генетического материала, способного размножаться в клетке и синтезирова
Значение групп крови для практики
Контроль достоверности происхождения животных.Одна из главных областей практического применения групп крови - контроль происхождения животных. Такое их использование вызвано тем,
Значение биохимического полиморфизма
Биохимические полиморфные системы белков используются для следующих целей:
1) изучения причин и динамики генотипической изменчивости, составляющей основу эволюционной генетики;
2
Генетический груз популяций
В ходе длительной эволюции животных наряду с полезными мутациями, подхватываемыми отбором, в популяциях или породах накопился определенный спектр генных и хромосомных мутаций. Каждое поколение по
Методы получения трансгенных животных
Трансгеноз - экспериментальный перенос генов, выделенных из определенного генома или искусственно синтезированных, в другой геном. Животные, в геном которых интегрируют чужеродные гены, наз
- Часть грудины. Грудная клетка. Особенности и возрастные изменения
- Харьковский патентно-компьютерный колледж Компьютерно патентный колледж
- Значение имени эрик, происхождение, характер и судьба имени эрик Имя эрих значение
- Магия чисел К чему снятся красные цветы на земле
- Страхование вкладов Порядок получения возмещения по вкладам
- Значение ростовцев яков иванович в краткой биографической энциклопедии
- Компот из вишни на зиму – только лучшие рецепты!